感谢你在百忙之中抽出时间阅读鲤脑垂体匀浆液和人绒毛膜促性腺激素混合注射对鳗鲡脑区促性腺激素释放激素和这篇文章。关于鲤脑垂体匀浆液和人绒毛膜促性腺激素混合注射对鳗鲡脑区促性腺激素释放激素和这篇文章的任何评论都可以告诉小编。你的每一个建议都是对小编辑的肯定和鼓舞。接下来让我们一起来了解鲤脑垂体匀浆液和人绒毛膜促性腺激素混合注射对鳗鲡脑区促性腺激素释放激素和。
汪小东林浩然中山大学生命科学学院广州510275谢刚中国水产科学研充院珠江水产研巧所,广州51旧80对雌暧脑区泡nRH_U则无屋著影响雌雄每次注射距H后l天,血清促性腺激素G旧急剧上巧。10天后逐渐下隆但雌在注射7次后1天血清曲水平仍然显著高于相应对照组。雄注射。6HGl4次后,血清睾丽浓度无思著变化;雌媛每次注射pEH后,血清睾邮浓度升高并直维持在较高水平。雌血清1巧_雌二醇浓度在注射46次期间与对照组无显著差异,注射7次后显著上升。
醇激素国家自然科学基金资助项目激素诱导缓娜卵母细胞*后成熟和产卵,39770101号和1典7高校博±点专项科研基金资助项目慢生殖活动的神经内分泌调节机理研究。
第作者简介;汪小东,男,巧折年7月生,博±。1吐020?818巨愚化邮弧拼4知61山客1£1客浊加。绅1合影比扣的繁素量愚含腺放素性释激促素醇膜激固毛腺类绒性性人促及和区素液脑激浆腺匀性体对促注射鱼类脑垂体匀浆液和人绒毛膜促性腺激素诱导性腺发育成熟的方法用于人工繁殖的研究,虽然不同研巧者所采用的脑垂体种类。催熟剂量。注射间隔时间等有所不同,但都取得较好的催熟效黑得到的成熟亲媛可诱导排卵和受精并得到幼曲3.然而用这种方法得到的幼苗在解出后10 ~15天大量死亡。其原因除了可能存在的生态条件不适外,不少学者提出用注射鱼类脑垂体匀浆液的方法诱导的媛成熟是异常发育134.遗憾的是,注射外源促性腺激素诱导媛性腺发育过程的内分泌调控机理的报道很少,口报道用注射蛙脑垂体诱导媛性腺发育成熟过程中类固醇激素含量的变化。本文采用口£和11混合注射的方法诱导性腺发育成熟,系统测定诱导媛性腺发育成熟过程中下丘脑脑垂体性腺轴中各个水平的激素含量变化,抖阐明外源促性腺激素诱导性腺发育成熟的内分泌调控机理。
1材料与方法1.1实验动物实验用媛放巧《祀火知曲为广东省珠江口渔民捕获的下海生殖涧游的鳄。雄鱼36尾,体长42体重120~260雌鱼拍币体长巧~78体重290~490护实验鱼蓄养于循环水族箱化暂养10~15天,然后逐步适应盐度为25~30的人工海水自然水温12~25°和光于19年11月至1996年5月、1996年11月至1997年5月间进行两批实验。
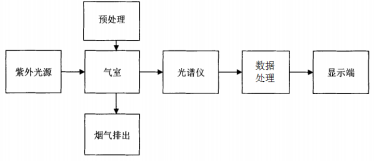
1.2狸脑垂体匀浆液的制备和注射方法狸脑垂体收自广州市农贸市场,保存在纯丙丽中。注射前用玻璃匀浆器磨碎,悬浮在Rin醉氏液中。注射剂量为:雌鱼每尾2.0g脑垂体100IU取G雄鱼减半,胸基部注间隔10天注射次。对照组慢注射虹巧氏液。
1.3样品收集和处理脑组织样品5次后1天[81:3.42抽。545]雌媛在注射前[幻:么化±0.、注射6次后1天[81:21.成±3.94]、注射10次后1天[81:50.78±6.41]取脑组织样品。取样时用碎冰将媛麻醉,切断脊椎,迅速取出脑组织置于冰浴的离屯、管中。脑组织分区参考文献[5]的方法分成H部分:A部分为端脑B部分包括间脑、中脑和下丘脑部分包括后脑和延髓。脑组织促性腺激素释放激素的抽提参考文献[6的方法稍做修改脑组织加入lL冰冷的2d/L古酸后,用超声波匀浆器匀浆,4°、巧000「/化离必1加化,取上清液在冷冻干燥仪上干燥,再重新溶解在测定用的盗酸缓冲液中10胃1电枯押7.4含化5路八,胆存于20尼待测抽和0含量。
血清样品雄媛共注射5~6次,分别于注射前和第1、2、3、4、5次注射后1天和10天取血清样品;雌鱼共注射片等逾哥猛律射前和第建7弓施盛谋遍占褚甜抓逼取血清愚巧呗在护次谭植/洁孟1巧小东等:取血清样品抖观察注射幻,611后血清激素含量的动态变化。取血清样品时,将媛用碎冰麻醉,用1血往射器从尾静脉取血,静置她后离必,血清胆存于2°[待测各种激素含量。取样时,解剖鱼体,计算成熟系数。
1.4激素含量的测定和数据处理媛脑区陆巧1含量的测定参考文献[7]建立的欧洲媛访3沿的测定方法,*小可测为20口客/血血清如1含量测定参考文献[巧,*低可测为化1唯/血性类固醇激素含量的测定参考文酬9]的方法,*低可测为巧9.数据抖平均值±标准误表示,两个平均值之间的差异显著性用8址加,81检验,两个抖上平均值之间的差异显著性用肥。孤8新复极差检验化05认为差异显暮1化1认为差异极显著。
2结果与分析2.1注射803对自曼局性腺发育成熟的影响两批实验共注射雄36尾,注射巧6~6次后,精巢战1大于4.0的有3尾,占实验鱼数的沿。335,对照组鱼件1均在化3.8之间。两批实验共注射雌性妨尾注射肥10 9~10次扁有钓39的实验鱼631达到30抖上,*大说1为化思%,对照组鱼的31小于5.0.
表明注射外源促性腺激素能较好地诱导雌雄媛性腺发育成熟。
注射061沿对媛脑区证狙含量的影响表l为注射距H诱导雄媛成熟过程中脑区GnRH含量的变化。注射前脑区各部分和总、1及沿脚11的含量都很低,八。8.焊鞑糠值暮棵挥邢灾钜臁W⑸浯次后脑区总。边化含量比注射前增加2.1倍,其中主要是6部分的加。注射5次后脑区磁01含量达到1低。
和。说±说。239其中主要是6和糠值脑黾印1砻髯⑸淦蓝0能促使雄媛脑前区八和BGnRH和脑后区化和GnRH?的含量加。
4.16pg,A部分和B部分都比注射4次后显著加P化01,部分GnRH含量在注射过程中较。脑区总、泡时1?11在注射前为泌。巧泌。9口W⑸浯次和5次后分别增加到进8.巧72.76注射PEHG诱导雄成熟过程中脑区GnRH含量的变化取样时间激素类型脑部分区脑区总含量注射前注射4次后注舶5次后注:平巧值标准误9脑组织,二8.
次后,n亿nRH总、含量加到注射前的1.6倍,其中A和B部分显著高于部分P化05.注ll0次后GnRH含量继续升高,脑区总、含量法到注射前的2.1倍,A和B部分均比注射6次时显著升高P化01,部分含量无显著变化P化05.脑区总GnRH?II含量在注射前为378.22±抑。5pg,是雄媛的典拍翁4溢PE巧Gf潘费虚盘揠趋部n蠢盟腧PEH巧滓加雌。net媛脑前区A和BGnRH的含量,而对GnRHII的含量无显著影响。
取样时间激素类型脑部分区脑区总含1注新前注射4次后注射5次后1口注;平巧值±标准误口皆脑组织,二8.
2.3注射电8对媛局血清0曲含量的影响雄媛血清Gff含量在每次注射a;H后l天均大于100ng/L.10天后下降至20~斯ng/址《对照组雄媛血清0出含量在整个实验过程中为化5~1.0£/址之间。
对照组雌媛血清QH含量在整个实验过程中无显著变化1.0~2.0ng/L.实验组雌媛每次注射色0后1天血清0出急剧上升30~70唯/血,注射4.5.6次10天后下降与相应对照组无思著差异£化05,注射7和8次后10天血清水平则仍然高于相应对照组图1.可见夕颇促性腺激素注入雌雄媛体内后,能立即引起血清3的含量的大幅度上升。
8次后雌媛血清3的的动态变化为:注射1天后显著上升,之后逐渐下隆至第10天时下降至与注射前无显著差异口。1图2.
注射后1天注巧后阳天口注射后1天0注巧后1天端虎注射4化注巧5次注射6次注射7化注射8妖巧昏1注射时间/d注射时巧/d注射后时巧/6 8次后雌血清激素含量的动态变化2.4注射汇8对血清性类固醇激素的影响雄媛注射0£1014次后血清睾丽浓度均低于793/加,与对照组相同。注射5次后1天血4~8次后1天和10天血清睾丽的浓度均显著高于相应对照组。6~化8雌/LパK化Olパ图l.表明注射PEHG后雌媛血清睾丽浓度直维持在较高且稳定的水平。
对照组雌血清1巧拉浓度为化2~0.3堪/加。实验组雌媛注射口6~6次后1天和1天血清1巧_62水平与对照组均无显著差异化05.注射7次和8次后1天,血清1邓_62含量水平分别达到a.62化21ng/L和l.抑化3汾ng/n山显著高于对照组和其他各次a化01,10天后分别下降至1.08±0.17皆址和1.08±0.化唯/血。但仍高于对照组,化版图1.
注射进£08次后雌媛血清睾醒和1巧62的动态变化丸注射时血清睾丽和巧62的浓度分别为4.55和1.0如£/血。注射后血清睾醒浓度在1~7天内略有上升5.20~5.64£/址3,10天后有所下降4.化ng/L,但均无显著差异P化版;血清l帘拉在注射后l天显著上升到l.烘ng/LF化01,3天和5天均保持在较高水平1.94和2.11ng/L,7天时下降为L30ng/L与注射时无显著差异化05图2.
3讨论3.1注射0屯1沿诱导媛局性腺发育成熟的效果注射幻;~6次后,有拟。33的雄媛031可达到4.0上,表明这种方法诱导媛精巢发育成熟具有良好的效果。在欧洲媛和日本,单独注射《就可诱导雄媛精巢发育成化11].
雌媛注射喃09~10次后,有妨。49的实验鱼31可达到30抖上,*大战1超过50,表明注射外源促性腺激素能有效地诱导雌媛卵巢发育成熟。
关于注射鱼类脑垂体诱导媛性腺发育成熟过程中所使用的种类和剂量各学者有所不同,欧洲学者注射狸鱼脑垂体剂量为2g/100gB.W,间隔2~3天注就约39次90天后部分雌媛卵巢发育达到成熟。日本学者注射蛙鱼脑垂体,剂量为2Wg/gB.W,间隔7天注射,约巧次后卵巢可达成熟口4].
我们采用趣鱼脑垂体和100混合注射,与王义强等使用的剂量相似1.从催熟效果来看,都可诱导媛卵巢发育成熟。因此,催熟剂的用量可能不是影响媛卵巢发育的重要因素。
二__日。穿/诉鹇巧3.2关于注射外源0肚诱导局性腺发育的机理注射鱼类脑垂体匀浆液和3诱导媛性腺发育的机理的研究报道还较少。本实验注射H后,脑区RnRH的含量随注射次数的增加而加,与欧洲媛的实验结果致n,进步证明了区班访出11的合成进而刺激脑垂体0出的合成来刺激性腺发育。雌在注射后,血清睾丽浓度加,睾丽可在脑和脑垂体水平通过正反馈作用刺激巧狂化11的合成队及脑垂体的的合成。在性未成熟虹自暮埋植或注射睾丽都能刺激GnRH和GtH的合成和释放[l2].在注射7次后lO天血清GtH仍在较高水平,可能与慢内源3的的合成和释放有关。雄注射R11后,血清睾丽浓度没有升高,可能睾丽被转化成其它性类固醇激素,如11_丽基睾丽再通过它起正反馈作用来刺激访很11和G出的合成和释放。注射PEHG后雌雄媛脑区GnRH含量的不同变化形式可能反应了雌雄脑区0站狙的不同分布。
雄精巢的发育成熟是在0出的作用下完成的。诱导内源的0的产生后,精巢可抖发育成熟。在切除脑垂体的欧洲雄,注射战同样可诱导精巢发育成熟,表明雄外源3衍与内源3在诱导精巢发育成熟时具有相同的生理作用。但在雌,单独注射成对卵巢发育毫无影响[3.表明在雌雄,性腺町受体的种类、结构、亲和性等都可能存在很大差异。
外源促性腺激素进入体内后的变化及其作用方式仍然不清楚。本实验结果表助注射〔后媛血清0出在注射后1天急剧升高,然后下降,再次注射后又是急剧上升。般来说,鱼类性腺发育是的缓慢而稳定增加的结果,的幅度较大的起落变化对媛性腺发育是否会产生重大影响还有待进步实验证明。血清睾丽浓度的增力日,表明外源促性腺激素刺激了睾生成但雌媛注射〔巧01~6次后,血清1审62的含量直没有加。离体研究也表强即使高浓度的睾丽为底慨再加入~450仙的离体卵母细胞生成啡62的能力仍然很低[4].说明在此期间卵巢缺乏将睾丽转变为l巧E2的芳化酶活性。在硬骨鱼类卵巢芳化酶是在GtH的作用下合成的,因此,夕h源促性腺激素可能对芳化酶的活性影响不大,注射7次后血清l本拉含量的加可能与内源QH的释放有关。然而,即使血清1巧_£2的含量在注射1~6次期间直较低与对照组无显著差异,卵巢却在不断地积累卵黄物质。我们可能未检测到1巧62的分泌高峰。外源促性腺激素进入鱼体后,由于受巧细胞上的受体类型、数量和亲和性等的影响,有可能在激素与受体结合后的转录或翻译过程产生变异,当然,此有待于进步的研究来检验。另外,鱼类脑垂体匀浆液中含有多种激素,各种激素的作用很难区分。由于在大多数硬骨鱼类存在两种促性腺激素泪的1和0的11,在蛙科鱼类的不同性腺发育时期,两种加1生理功能分化较为明强巧。*近,¥8证明在媛同样存在两种31比但它们各自在性腺不同发育阶段的生理功能仍不清楚141.在欧洲注射纯化的鱼3出11可抖促进性腺发育13,该方面的工作仍有待于更深入的研究。脑垂体匀浆液中其它种类的激素,如生长激素。促现素、促甲状腺激素、促肾上腺皮质激素等对亲的物质代谢和卵巢发育所产生的影响也值得进步研王义强,赵长蕾施正峰等。河蒺人工繁殖的初步研究。水产学报,巧8042:147~ 2期汪小东等;林浩然,张梅丽,张素敏等。慢繁殖生物学研究人工催熟过程中下海慢娜的分泌活动、性腺发育状况和脑垂体世细胞的超显微结构。水生生物学报,198了,328驴础化如1.2过8浊1典1,81说